



Podcast su il Diario di Anna Frank
Podcast su il Diario di Anna Frank: file audio scaricabile gratuitamente sul libro che narra la persecuzione nazista con gli occhi di una ragazza ebrea

Podcast su il Diario di Anna Frank: file audio scaricabile gratuitamente sul libro che narra la persecuzione nazista con gli occhi di una ragazza ebrea

Podcast sul Manifesto degli intellettuali fascisti e l'Antimanifesto: file audio scaricabile gratuitamente sui due documenti di Giovanni Gentile e Bendetto Croce

Podcast su Sparta e Atene: somiglianze e differenze tra due pòleis diverse tra loro, alleate e nemiche nel corso della storia dell'antica Grecia

Podcast sulla Giornata della Terra: file audio scaricabile gratuitamente sulla giornata che ogni 22 aprile sensibilizza sulle risorse del pianeta

Podcast su Alan Turing e la decodifica di Enigma: file audio scaricabile gratuitamente sulla biografia dello scienziato e il suo ruolo nella Seconda guerra mondiale

Podcast sulla revisione del testo: file audio scaricabile gratuitamente sulle tecniche per fare una revisione di un testo

Podcast sull'intelligenza emotiva: file audio scaricabile sul tipo di intelligenza che serve a comprendere le emozioni e sviluppare relazioni positive ed efficaci

Podcast sui sovrani più crudeli della storia passata: file audio scaricabile gratuitamente sugli imperatori, i re, gli zar e i papi passati alla storia per i loro atti efferati

Podcast sull'overthinking: file audio scaricabile sul disturbo che causa lo sviluppo costante di pensieri intrusivi e rimuginio

Podcast sulla FOMO: file audio scaricabile gratuitamente sulle cause e conseguenze della Fear of Missing Out. Cos'è e come affrontarla

Podcast sul testo espositivo: file audio scaricabile gratuitamente su come si fa un testo espositivo e quali sono le sue caratteristiche

Podcast sulle origini del Natale: da dove nasce la tradizione occidentale e quali sono i riferimenti sociali e culturali precedenti

Podcast sulla separazione dei poteri: file audio scaricabile gratuitamente sul pensiero di Montesquieu e sulla storia precedente

Podcast su diario, lettera e autobiografia: file audio scaricabile gratuitamente sulla differenza e le somiglianze fra i generi letterari

Podcast sui pianeti del Sistema solare: file audio scaricabile gratuitamente su somiglianze e differenze fra i pianeti del sistema solare

Podcast sull'Unità d'Italia: file audio scaricabile gratuitamente sugli eventi e i protagonisti che hanno portato alla creazione dell'Italia unita nel 1861

Podcast su Elisabetta II: file audio scaricabile gratuitamente sulla regina dei record, che ha regnato per 70 anni in UK e nel Commonwealth

Podcast sul mito di Apollo e Dafne: file audio scaricabile gratuitamente con il racconto del mito e spiegazione del suo significato simbolico

Podcast su Io non ho paura di Niccolò Ammaniti: file audio scaricabile gratuitamente sul romanzo del 2001, con trama, personaggi e significato

Podcast sulla Favola di Amore e Psiche: file audio scaricabile gratuitamente sul mito del figlio di Venere e della fanciulla che superò prove per restargli accanto

Podcast sulla nascita della festa di halloween: file audio scaricabile gratuitamente sulle origini della festa e del nome

Podcast sulla Gioconda di Leonardo da Vinci: file audio scaricabile gratuitamente con descrizione del dipinto e significato dell'opera

Podcast su Il piccolo principe. File audio scaricabile gratuitamente sul romanzo più famoso del francese Antoinde De Saint-Exupéry

Podcast su Frankenstein di Mary Shelley: file audio scaricabile gratuitamente su trama e significato del romanzo gotico più famoso di sempre

Podcast su Ciaula scopre la luna di Luigi Pirandello: file audio scaricabile gratuitamente su trama e significato della novella dello scrittore siciliano

Podcast su come si fa una scheda libro: file audio scaricabile gratuitamente sulle tecniche per creare una perfetta scheda libro

Podcast su eroe e antieroe da Omero al Novecento: file audio scaricabile gratuitamente su come cambia il concetto di eroe e antieroe in letteratura

Podcast su 1984 di George Orwell: file audio scaricabile gratuitamente su trama e analisi del romanzo di fantascienza distopica dello scrittore inglese

Podcast sull'Esposizione universale: storia e caratteristiche dei principali Expo che si sono susseguiti in Europa dalle origini a oggi

Podcast su Leonardo da Vinci poeta e letterato: file audio scaricabile gratuitamente sulla produzione letteraria del grande genio toscano

Podcast su Emilio Salgari e il romanzo d'avventura: file audio scaricabile sullo scrittore di romanzi ambientati in luoghi esotici e lontani

Podcast sul capitolo 1 dei Promessi sposi, il celebre romanzo scritto da Alessandro Manzoni. Audio lezione scaricabile gratuitamente

Podcast sulla storia dell'alfabetizzazione in Italia: file audio scaricabile gratuitamente sulle riforme della scuola e l'alfabetizzazione del popolo italiano

Podcast sul canone di bellezza nella storia: file audio scaricabile gratuitamente sul modo in cui è cambiato nei secoli il concetto di bellezza

Podcast su Il deserto dei Tartari di Dino Buzzati: file audio scaricabile gratuitamente sul celebre libro dello scrittore e giornalista italiano
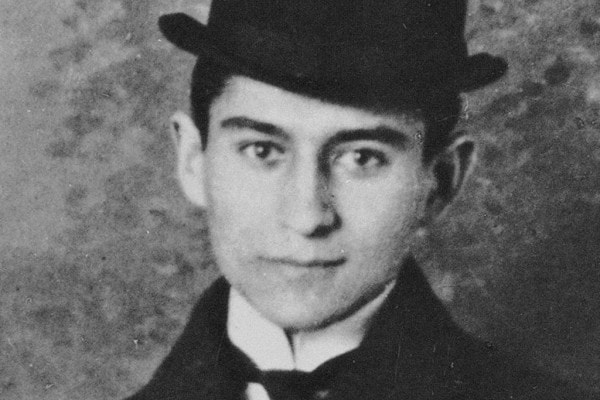
Podcast su Franz Kafka: file audio scaricabile gratuitamente su vita e opere principali dell'autore di la Metamorfosi e Lettera al padre

Podcast su Fausto Coppi, il grande campione del ciclismo italiano del dopoguerra. Vita, tappe fondamentali, storia privata del ciclista

Podcast su cosa è cambiato nel cinema dalla nascita ai giorni nostri: dai film muti fino ai moderni blockbuster. La storia del cinema in breve

Podcast su Tutankhamon e la scoperta della sua tomba: file audio scaricabile gratuitamente sulla storia del Faraone bambino e la sua ricca tomba

Podcast sulla Rivoluzione francese: file audio scaricabile gratuitamente sulla caduta dell'ancien régime e l'avvento dei rivoluzionari in Francia

Podcast su Maria Montessori, pedagogista, educatrice e neuropsichiatra infantile. File audio scaricabile sulla sua vita e la sua teoria pedagogica

Podcast su Pier Paolo Pasolini: file audio scaricabile gratuitamente sullo scrittore, regista, poeta e drammaturgo italiano ucciso ad Ostia nel 1975

Podcast su Edvard Munch e l'Urlo: file audio scaricabile gratuitamente sulla biografia del pittore scandinavo e la sua opera più famosa, Il Grido

Podcast su Dante Alighieri: file audio scaricabile gratuitamente sul padre della lingua italiana, una delle "Tre corone" insieme a Petrarca e Boccaccio

Podcast su Cesare Beccaria: file audio scaricabile gratuitamente su vita e opere del giurista milanese autore di Dei delitti e delle pene

Podcast su Francesco Petrarca: file audio scaricabile gratuitamente su una delle "Tre corone" della lingua italiana del Trecento

Podcast sull'emigrazione europea negli Stati Uniti: file audio scaricabile gratuitamente sull'emigrazione europea nel Nuovo Continente dall'800

Podcast sulla questione della lingua italiana: file audio scaricabile gratuitamente sulla nascita del volgare e il suo ruolo nei secoli

Podcast su Rustico e Alibech, la decima novella della terza giornata del Decameron di Boccaccio. File audio scaricabile gratuitamente
Podcast su Calandrino e l'elitropia, terza novella dell'ottava giornata del Decameron di Boccaccio. File audio scaricabile gratuitamente

Podcast su Artemisia Gentileschi: file audio scaricabile gratuitamente sulla pittrice, la sua arte potente ed espressiva e la sua biografia tormentata
Podcast su Peronella, seconda novella della settima giornata del Decameron di Boccaccio. File audio scaricabile gratuitamente

Podcast sulla stregoneria: file audio scaricabile gratuitamente sulla storia della stregoneria e della caccia alle streghe, dagli inizi al '700
Podcast su Guido Cavalcanti, nona novella della sesta giornata del Decameron di Boccaccio.File audio scaricabile gratuitamente

Podcast sul Dolce Stilnovo: file audio scaricabile gratuitamente sulla poesia amorosa del Trecento, focalizzata sulla donna-angelo

Podcast sulla Guerra Fredda: file audio scaricabile gratuitamente sul conflitto che vide protagonisti USA e URSS dopo la Seconda guerra mondiale

File audio con spiegazione sulla storia, dalla nascita alla caduta, del Muro di Berlino. Podcast da ascoltare, scaricare e archiviare gratis
Podcast su Frate Cipolla, l'ultima novella della sesta giornata del Decameron di Boccaccio. File audio scaricabile gratuitamente

Podcast sull'età comunale in Italia: file audio scaricabile gratuitamente sull'Italia dei Comuni nel Medioevo. Cosa sono e come funzionano
Podcast su Nastagio degli Onesti, ottava novella della quinta giornata del Decameron di Boccaccio. File audio scaricabile gratuitamente